








| Map of Danish marine waters | ||
| Eelgrass in coastal waters | ||
| Long-term changes in eelgrass distribution | ||
| Recent inter-annual fluctuations in eelgrass distribution | ||
| Short-lived nuissance macroalgae | ||
| Macroalgae on reefs in open waters | ||
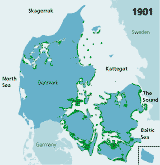
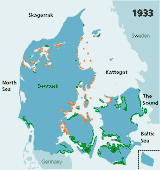
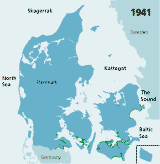
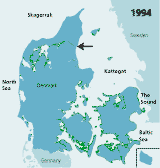
| Figures 2.32 Map of eelgrass area distribution in Danish coastal waters in 1901 (redrawn after Petersen 1901), 1933 (redrawn after Blegvad 1935), 1941 (redrawn after Lund 1941) and 1994 (coarse map based on visual examination of aerial photos and data from the national Danish monitoring programme, produced by Jens Sund Laursen). Green areas indicate healthy eelgrass while orange areas (on the 1933 map) indicate where eelgrass was affected by the wasting disease but still present in 1933. The arrow shows the location of Limfjorden. (Boström et al. 2003). |
| 1901 |
 |
| 1933 |
 |
| 1941 |
 |
| 1994 |
 |
Figures
2.33 |
| Open sea coast |
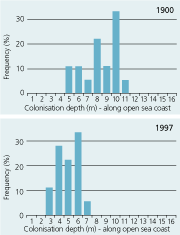 |
| Estuaries |
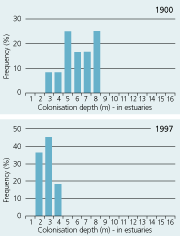 |
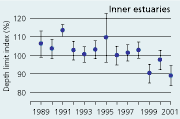
| Figures
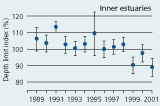
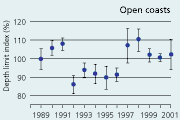
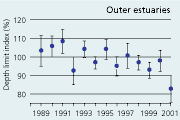
2.34 Index of maximal eelgrass colonisation depths (± s.e.) during the period 19892001 along open coasts, outer estuaries and in inner estuaries. Index values represent colonisation depths of a given year relative to average colonisation depths of the period 1989 2001 (Ærtebjerg et al. 2002). |
| Open coasts |
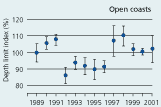 |
| Outer estuaries |
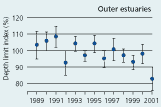 |
| Inner estuaries |
 |
| Figure 2.35 |
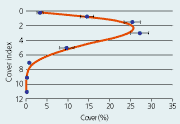 |
| Eelgrass cover as a function of depth. Data represent average cover of 276 depth gradients monitored under the national Danish Aquatic Monitoring and Assessment Programme in 1994. |
| Figures
2.36 Cover index of loose lying nuisance macroalgal species in the inner parts of estuaries during the period 19932001. Data represent the depth intervals 01 m, 12 m and 24 m. The cover for each depth interval in a given year has been indexed relative to average cover of the depth interval during the period 19932001 (Ærtebjerg et al. 2002). |
| 0-1 meter |
 |
| 1-2 meter |
 |
| 2-4 meter |
 |
| Figure 2.37 |
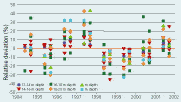 |
| The relative deviation in macroalgal cover in relation to mean values for individual stations and time of investigation. Deviations are given in 2 meter depth intervals. Stations where algal abundance was limited by sea urchin grazing are left out. |
Submerged aquatic vegetation
Eelgrass (Zostera marina) is the most widely distributed marine angiosperm in shallow Danish coastal waters. On hard substrates the vegetation is dominated by macroalgae. Growth of short-lived nuisance species of macroalgae is a problem in inner parts of some estuaries.
EELGRASS IN COASTAL WATERS
Eelgrass and seagrasses in general often represent a large standing biomass
and a high primary production that influence the overall functioning of
coastal ecosystems. Seagrasses enhance biodiversity by providing habitat
and shelter for many species; they are nursery and foraging areas for
commercially important species of fish; they improve water quality by
reducing particle loads and absorbing dissolved nutrients; they stabilise
sediments and influence global carbon and nutrient cycling (see Hemminga
& Duarte 2000). The many ecosystem services provided by eelgrasses
have created a concern for the response of this species to eutrophication.
This concern has been intensified over the last few decades where large-scale
reductions in seagrass meadows have been reported in response to eutrophication
from many areas (see Short & Wyllie-Echeverria
1996).
The following paragraphs summarise the development of Danish eelgrass meadows during the last century. The presentation is based on early reports on eelgrass distribution covering the period 1900-1940, aerial photos from the archives of KMS (The National Survey and Cadastre Agency) covering the period 1945–1990s and data from the national Danish monitoring programme on marine vegetation covering the period 1989–2001.
LONG-TERM CHANGES IN EELGRASS DISTRIBUTION
In Denmark, records of eelgrass distribution date back to around 1900,
and provide a unique opportunity to describe long-term changes. In 1900,
eelgrass was widely distributed in Danish coastal waters, and covered
approximately 6726 km2 or 1/7 of all Danish marine waters (Petersen
1901, 1914) (Figure
2.32). The standing crop made up almost 1 kg dw m-2 in
the densest stands and the total annual eelgrass production was estimated
at 8 million tonnes dry weight (Petersen
1914). In the 1930s, the world wide wasting disease substantially
reduced eelgrass populations, especially in north-west Denmark where salinity
is highest (Blegvad 1935). In
1941, eelgrass covered only 7% of the formerly vegetated areas and occurred
only in the southern, most brackish
waters and in the low saline inner parts of Danish estuaries
(Lund 1941, Rasmussen
1977)(Figure 2.32). No national monitoring existed
between 1941 and 1989 but analyses of aerial photos from the period 1945–1990s,
reveal an initial time lag of more than a decade before substantial recolonisation
of the shallow eelgrass populations began. The photos also show that large
populations had recovered in the 1960s (Frederiksen
et al. in press.).
Today eelgrass again occurs along most Danish coasts (Figure 2.32) but has not reached the former area extension (Olesen 1993, Boström et al. 2003). Comparisons of eelgrass area distribution in two large regions, Øresund and Limfjorden, in 1900 and in the 1990s, suggest that the present distribution area of eelgrass in Danish coastal waters constitutes approximately 20–25% of that in 1900. The area distribution of eelgrass in Limfjorden was estimated at 345 km2 in 1900 (Ostenfeld 1908) and only at 84 km2 in 1994 (based on aerial photography data from the Limfjord counties). In Øresund, eelgrass covered about 705 km2 in 1900 (Ostenfeld 1908) and only about 146 km2 in 1996–2000 (Krause-Jensen et al. 2001). Differences in methodology may, however, influence these comparisons.
The large reduction in area distribution of Danish eelgrass meadows is partly attributed to loss of deep populations. In 1900, colonisation depths averaged 5–6 m in estuaries, and 7–8 m in open waters while in the 1990s, colonisation depths were reduced about 50% to 2-3 m in estuaries and 4–5 m in open waters (Figure 2.33) (Boström et al. 2003). The deep populations are most likely lost as a consequence of eutrophication. Increased nitrogen concentrations stimulate phytoplankton growth and thereby reduce the transparency of the water column and restrict the colonisation depth (Nielsen et al. 2002).
RECENT INTER-ANNUAL FLUCTUATIONS IN EELGRASS DISTRIBUTION
Since 1989, the Danish Aquatic Monitoring and Assessment Programme has
included annual surveys of colonisation depth and cover of eelgrass along
depth gradients in a wide range of estuaries and coastal waters.
The colonisation depth of eelgrass reflects differences in water quality and physical setting along estuarine gradients. In the inner parts of estuaries, the annual mean colonisation depth ranged between 2.9 and 3.5 m, in outer parts between 3.3 and 4.3 m, and along open coasts between 4.7 and 5.8 m during the period 1989–2001 (Figure 2.34). Eelgrass colonisation depth shows no significant trend between 1989 and 2001 and does not reflect the sligh amelioration of water clarity observed through the same period (Ærtebjerg et al. 2002). Other regulating factors may blur the relationship between light and colonisation depth so that more marked changes in light climate are needed before colonisation depths increase. For example, eelgrass suddenly disappeared from several sites during the warm summers of 1992 and 1994 possibly due to combined exposure to anoxia, sulphide and extreme temperature (Goodman et al. 1995, Terrados et al. 1999).
Eelgrass displays a bell-shaped distribution pattern along the depth gradient with maximum abundance at intermediate depth and lower abundance in shallow and deep water (Figure 2.35). Exposure, desiccation and ice-scour act to reduce eelgrass abundance in shallow water and render the populations extremely dynamic and unpredictable. In the deeper, more protected waters, reductions in eelgrass abundance towards the lower depth limit correlate with light attenuation (Sand-Jensen et al. 1997, Krause-Jensen et al. 2000) and are therefore more directly coupled to changes in eutrophication. The period 1989–2001 showed no significant trend in eelgrass cover at water depths above 2 m, but the cover of shallow populations was significantly reduced in inner estuaries and along open coasts (Henriksen et al. 2001). We have found no obvious explanation for this pattern.
In conclusion, eelgrass responds to several types of disturbances: changes in energy input (light), physical disturbances (e.g. wind, waves, extreme temperature, ice), chemical disturbances (e.g. anoxia, sulphide) and biological disturbances (e.g. the wasting disease). When eelgrass is used as a monitoring parameter to reflect changes in light climate due to eutrophication we should therefore be aware that other factors may affect the response. As the intensity of physical disturbances decline with depth, eelgrass colonisation depth and abundance from intermediate depths towards deeper waters are therefore likely to be better response parameters to eutrophication than the abundance of shallow populations.
SHORT-LIVED NUISANCE MACROALGAE
Short-lived nuisance macroalgae are favoured by large supplies of nutrients
(Pedersen 1995). Together with
macroalgal species numbers and dominance patterns, the cover of nuisance
species is therefore used as an indicator of the state of macroalgae in
coastal areas in the Danish Aquatic Monitoring and Assessment Programme.
Information on these algae is relatively scarse and analyses of trends are therefore only possible for the inner part of estuaries at depth intervals of 0–1, 1-2 and 2–4 meter. During the monitoring period (1993–2001) average relative cover varied from 1–20% at 0–1 meter depth, 1–25 % at 1–2 meter depth and 0–20% at 2–4 meter depth. There were no significant changes during the period 1993 to 2001 (Kendalls-tau, p > 0,05) (see figure 2.36).
MACROALGAE ON REEFS IN OPEN WATERS
As with seagrasses the macroalgae on the reefs in the open waters enhance
biodiversity by providing habitat and shelter for many species. Furthermore
they constitute a great part of the bio-diversity in marine vegetation.
The vegetation in the open waters consists of a multilayer of red and
brown algal vegetation at water depths of 10–12 meters. At depths
greater than 12–14 m total cover of upright algae decreases to a
single layer with coverage less than 100% and cover decreases further
with increased depth. In addition to upright forms, crust forming macroalgae
cover stones and shells. The cover of crust forming algae is large, even
at 24–25 meters depth. Macroalgal vegetation is monitored at 9 stone-reefs
and rocky bottom areas evenly distributed in the Kattegat, with one station
located in the northern Belt Sea. The total relative cover of the upright
forms is described along with specific cover of individual species including
crust forming algae.
It has been shown that a good empirical correlation exists between the
supply of the inorganic nutrients (nitrogen and phosphorus in freshwater
inputs) and total cover of macroalgae at deep stations in the Kattegat
during the period 1994 to 2000 (Henriksen
et al. 2001). This means that high supplies of inorganic nutrients
or freshwater leads to a reduced development of benthic vegetation. Exceptions
are stations with an intense grazing pressure from the sea urchin Strongylocentrotus
droebachiensis.
There is a corresponding significant correlation between the cover of
benthic vegetation and the pelagic parameters, Secchi depth and chlorophyll
concentration. This supports the hypothesis that the supply of nutrients
influences light penetration to the bottom and thereby the environmental
quality of the individual reef locations.
The total cover of the upright vegetation tended to increase in 2001 relative to the mean of the period 1994–2001 (Table 2.9). In general, algal coverage was low in years with relatively high runoff and high in years of low runoff. Accordingly there was no overall trend in the cover of macroalgae at the monitored reef stations (Figure 2.37).
Table 2.9
Macroalgal cover at a number of stonereefs in Kattegat
relative to the average cover in the period 1994–2001.
![]() means that
the majority of examined stations has a more developed vegetation cover.
means that
the majority of examined stations has a more developed vegetation cover.
![]() means that the majority of examined stations has a less developed vegetation
cover.
means that the majority of examined stations has a less developed vegetation
cover.
![]()
![]() means that there is an equal number of stations with a less respectively
more developed vegetation cover.
means that there is an equal number of stations with a less respectively
more developed vegetation cover.
A signtest shows whether cover has increased or decreased relative to
the mean for the period 1994-2001. * = P < 5%, ** = P < 1%, ***
= P < 0,1%.
| Year |
Month |
Number of observations |
Macroalgae coverage with reference to average coverage 1994-2001 |
P |
| 1994
|
June
August |
4 11 |
– – |
|
| 1995 |
June August |
11 12 |
– – |
|
| 1996 |
June August |
9 9 |
– * |
|
| 1997 | June August |
11 11 |
*** *** |
|
| 1998 | June August |
10 10 |
* ** |
|
| 1999 | June August |
10 11 |
* *** |
|
| 2000 | June August |
10 12 |
– – |
|
| 2001 | June August |
11 11 |
– – |
| | Top | |
| Next
| |